Nguyen ,
Shapiro-Ilan , Stuart, McCoy
, James & Adams,
2004
Male: (Fig.
1)
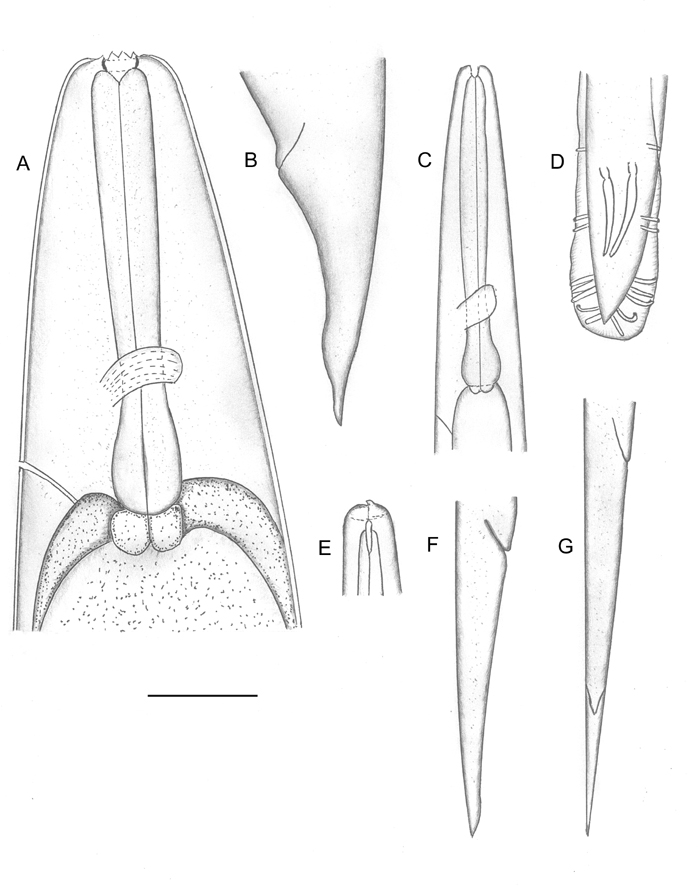
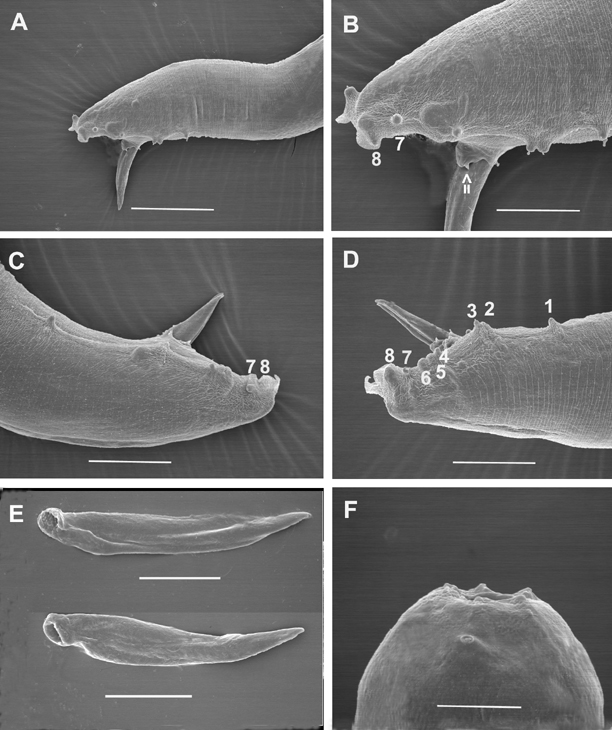
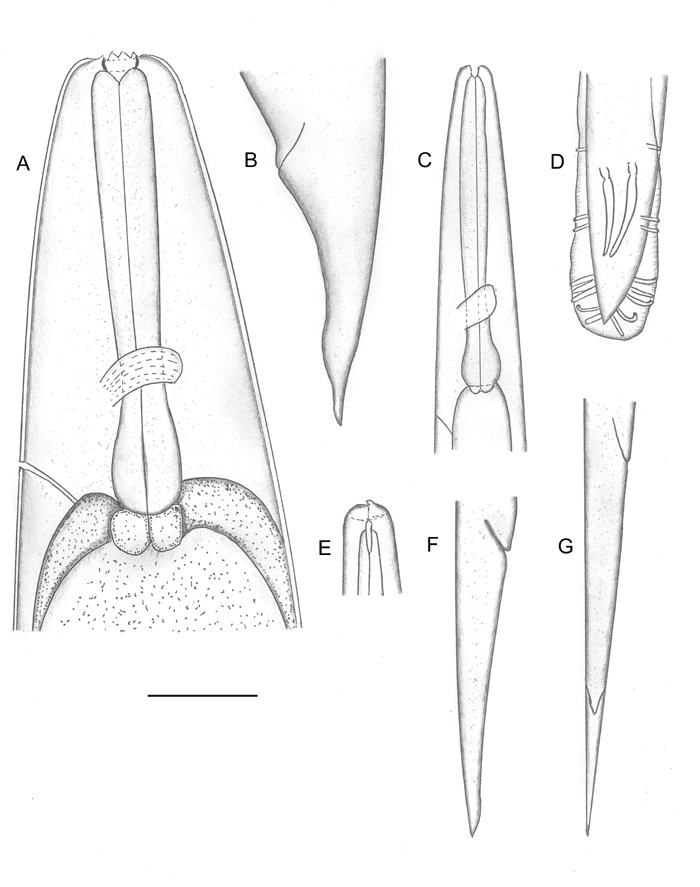
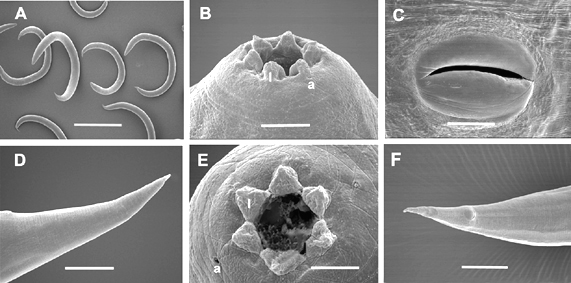
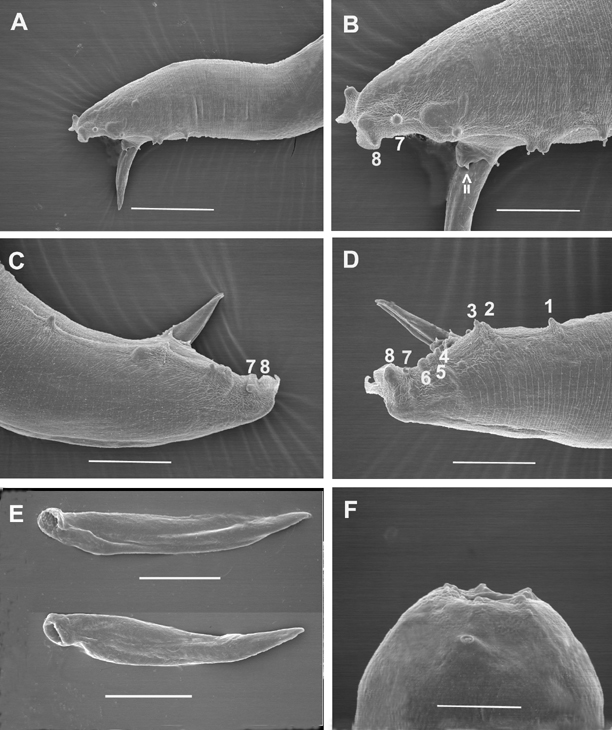
Body curved ventrally when heat killed. Head truncate, sometimes slightly swollen. Labial papillae 6, cephalic papillae not observed. Amphid prominent (Fig. 2F). Stoma with sclerotized, rod- or barrel-shaped cheilorhabdions; other rhabdions not distinguishable forming a funnel-shaped structure below cheilorhabdions. Pharynx with cylindrical corpus, metacorpus sometimes slightly enlarged. Nerve ring surrounding isthmus just anterior to basal bulb. Basal bulb with reduced valve. Cardia present, protruding into intestine. Excretory pore usually posterior to basal bulb. Testis monorchic, reflexed; distance from end of basal bulb to testis flexure variable. Vas deferens well developed. Spicules paired, separate, rostrum absent, ventral side usually flattened. Gubernaculum slightly curved ventrally. Bursa peloderan. Seventy percent (28/40) of observed males with 8 pairs of bursal papillae (Fig. 2). Three pairs anterior to cloaca and 5 pairs posterior to cloaca. From anterior to posterior, pair 1 well anterior, its tips reaching beyond bursal rim; pairs 2 and 3 in a group, immediately anterior to cloaca, also reaching beyond bursal rim. Pairs 4, 5 and 6 forming a group, situated just posterior to cloaca, pair 4 usually curved outward (laterally). The number of pairs from 1-6 are constant. Pairs 7 and 8 forming the terminal group of papillae (Fig. 3) at the end of bursa (Fig. 2). Pair 7 always curved outward (laterally). About 30% of males do not have 2 pairs in this last group of papillae. Instead, they have either three papillae on one side and two or one papillae on the other side, or only one pair. In addition to bursal papillae, a pair of smaller papillae was observed on the posterior edge of cloaca (Fig. 2B-D). Tail conoid, slightly curved ventrally.
Female
(Fig. 1)
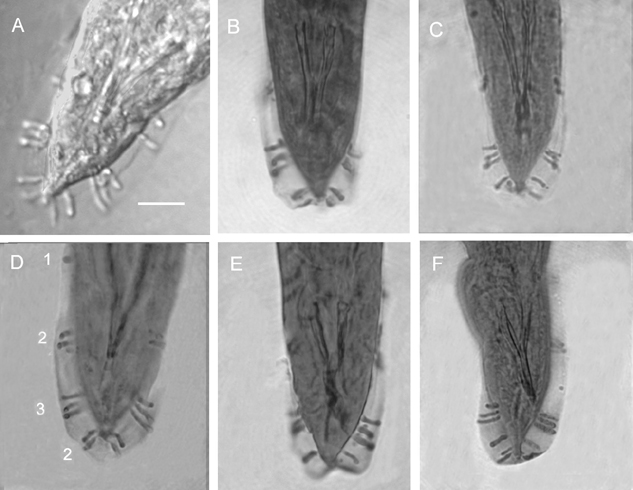
Hermaphroditic female C-shaped in fixative, body robust, always with many eggs in young females and many eggs and juveniles in mature females. Cuticle appearing smooth under light microscope but finely annulated with SEM. Head region tapering anteriorly. Labial region with 6 prominent lips, each lip with a labial papilla on top (Fig. 4) Cephalic papillae not observed with SEM. In face view, mouth hexagonal in shape as in amphimictic female in Fig. 5E. Amphid small. Stoma with refractile cheilorhabdions appearing as a circle below lips. Posterior part of stoma funnel-shaped, enclosed by anterior part of pharynx. Pharynx with cylindrical corpus. Isthmus present. Nerve ring at middle of isthmus. Basal bulb prominent with inconspicuous valve, but lumen of pharynx in basal bulb well sclerotized. Cardia present. Gonads didelphic, amphidelphic. Vulva with a transverse slit, located on a protruding area, anterior to mid-body. In ventral view, vulva top smooth, elliptical. This structure was observed on at least 20 specimens under SEM. Vagina short. Tail longer than anal body diameter, conoid with pointed terminus. Sometimes, near the end, tail widens then narrows down to a pointed tip. Post-anal swelling present. Phasmid inconspicuous. Amphimictic female similar to hermaphroditic female but smaller.
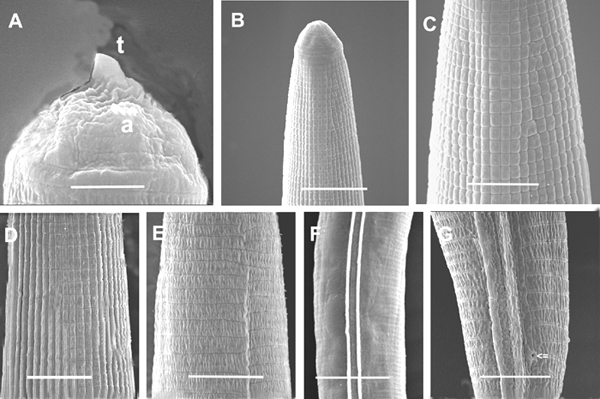
Ensheathed infective juvenile (third stage inside second stage cuticle) with body length close to that of H. bacteriophora and H. indica. Labial region with 7 annules, without longitudinal incisures. Anterior part of body (about 3-5 body diameters from anterior end) with tessellate pattern. Posterior part with longitudinal ridges. Tail long, pointed. Exsheathed infective juvenile body annulated, without longitudinal ridges. Labial region with a dorsal tooth, labial and cephalic papillae absent. Amphid prominent. Excretory pore posterior to nerve ring but anterior to base of pharynx. Pharynx typical for Heterorhabditis. Lateral field beginning anteriorly with one ridge (Fig. 5). A short distance posteriorly (2-3 body diameters), one more ridge appears to make 2 ridges in lateral field. This pattern is unchanged for the rest of the body. Phasmid near mid tail, pore-like, prominent, just dorsal to lateral field (ventral to lateral field in Steinernema: Nguyen & Smart, 1993, Nguyen & Adams, 2003). The presence and location of phasmids have not been reported for other species of Heterorhabditis. Tail elongate conoid with bluntly pointed terminus. Tail length without sheath is about 67% of tail with sheath.
Sorghum field, approximately 4 miles south of Nuevo Progresso, in the northern part of the state of Tamaulipas, Mexico.
Holotype (male), allotype (female) and paratype (hermaphrodites, females, male, and infective juveniles in 3% formalin) isolated from haemocoel of G. mellonella deposited in the United States Department of Agriculture Nematode Collection (USDANC), Beltsville, Maryland.
Morphologically, the new species is most similar to H. bacteriophora, H. brevicaudis, and H. indica, and can be distinguished (diagnosed) from these species mainly by male and female characters. For males of H. mexicana n. sp., 70% have 8 pairs of bursal papillae (Fig. 2,3) compared to nine or variable number in other species (Fig. 6). GS values are higher than that of H. bacteriophora, H. brevicaudis, and H. indica (Table 2), and the SW value is lower than that of all other species. For females, the vulval pattern of the new species (Fig. 4C) is quite different from that of the other morphologically similar species, H. bacteriophora, and H. indica (Nguyen & Smart, 1998). This pattern resembles that of H. zealandica as indicated by Adams and Nguyen (2002). The vulval pattern of H. brevicaudis could not be obtained for comparison. The new species can be distinguished from H. megidis, H. zealandica, and H. marelatus by the number of bursal papillae (Fig. 6A-C).
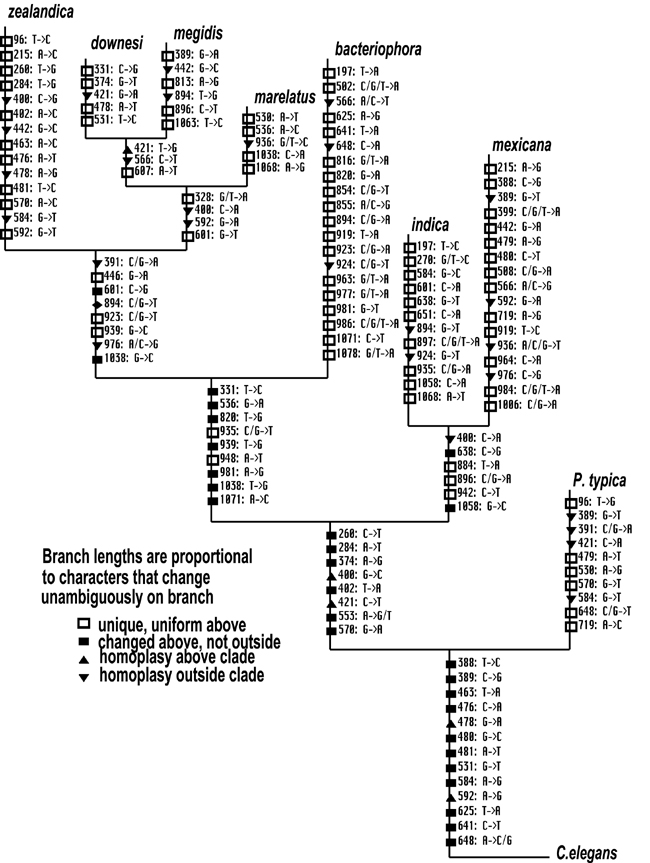
Parsimony, likelihood and distance based treebuilding approaches produced a single, concordant phylogenetic hypothesis for the genus Heterorhabditis (Fig. 7). H. mexicana n. sp. has evolved 13 autapomorphic nucleotide character states, and differs from its sister taxon H. indica at 113 aligned positions of the complete ITS rDNA array (Fig. 7). Although the present study nearly doubles the amount of molecular data of previous analyses of the genus (Adams et al., 1998), relationships among the nominal taxa remain unchanged. While branch support (via bootstrapping) is much improved for the parsimony analyses, certain nodes remain weak in likelihood and distance analyses. Particularly weak are nodes involving the placement of H. zealandica relative to H. marelatus, H. downesi, and H. megidis, and for the position of H. bacteriophora relative to H. indica and H. mexicana n. sp. (data not shown). Mapping of these characters for the purpose of informing species delimitation assumes that the autapomorphic characters we have identified are fixed among populations of H. mexicana n. sp. Certainly this assumption is violated for at least few nucleotide positions, which could be expected to vary among different populations (Adams et al., 1998; Powers et al., 1997). Although ITS rDNA sequences of H. mexicana n. sp. were sampled from other populations (also from Tamaulipas) and no DNA sequence heterogeneity was found (data not shown), we did not approach the requisite sample sizes to infer character fixations with statistical confidence (Sites & Crandall, 1997). However, we feel that our suite of unique morphological characters combined with the numerous molecular autapomorphies we uncovered are sufficient to identify cladogenesis and delimit H. mexicana n. sp. as a single lineage evolving independently from its sister taxon.
Reference
Nguyen, K. B.
, Shapiro-Ilan, D. I., Robin
J. Stuart, R. J,Clay W. McCoy, C. W., Rosalind R. James, R. J, & Adams, B. J. 2004. Heterorhabditis mexicana n. sp.
(Rhabditida: Heterorhabditidae) from Tamaulipas, Mexico, and morphological
studies of the bursa of Heterorhabditis spp. Nematology 6:231-244.
_____________________________________________
This website is
maintained by Khuong B. Nguyen
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}